Mangrove
La mangrove est un écotone, soit un espace de transition entre des écosystèmes terrestres et des écosystèmes marins. Ainsi, la mangrove est soumise aussi bien à la dynamique côtière (courants de marée, flot, jusant, etc.) qu’aux différents processus d’origine terrestre (flux d’eau douce charriant nutriments et sédiments). Ce type d’écosystème particulier peut évoluer sur des substrats sablo-vaseux, comme c’est le cas la plupart du temps, ou sur des substrats rocheux.


Sur les substrats vaseux, les palétuviers doivent faire face à un sol meuble, salé et privé d’oxygène car saturé en eau même à marée basse. Ils ont ainsi développé d’étonnantes facultés d’adaptation. Au niveau racinaire, le genre Rhizophora se caractérise par le développement de racines échasses en forme d’arceaux qui assurent un ancrage efficace. Il en est de même pour le genre Bruguiera mais ces dernières sont plus ramassées à la base du tronc (PASKOFF, 2004). Les espèces d’Avicennia et de Sonneratia ont développées des pneumatophores, soit des appendices d’allures stalagmitiques leur permettant de récupérer de l’O2 (PASKOFF, 2004). En effet, le sol hydromorphe des mangroves est dit « anoxique » car pauvre en dioxygène (structure du sol non poreuse).


La viviparité chez les espèces du genre Rhizophora est également un signe d’adaptation aux conditions difficiles du milieu : lorsqu’elles tombent au niveau du sol, les graines déjà germées pourront plus facilement s’implanter lors de leur dispersion soumise aux conditions hydrodynamiques. Les palétuviers sont halophytes, c'est-à-dire qu’ils sont conçus pour supporter de fortes salinités. Leur pression osmotique est élevée. Rhizophora possède un mécanisme de filtrage dans son système racinaire qui empêche le passage du sel. Celui-ci pénètre dans Avicennia mais il est éliminé par excrétion et cristallise à la surface des feuilles (PASKOFF, 2004).
Une soixantaine d’espèces de palétuviers sont présentes dans le monde mais il existe des disparités entre les trois bassins océaniques que sont l’Atlantique, l’océan Indien et le Pacifique. Les espèces sont plus nombreuses dans les mangroves des rivages de l’océan Indien et l’océan Pacifique occidental que dans celles qui bordent les côtes de l’Afrique de l’Ouest et des deux Amériques (PASKOFF, 2004). Environ 50% des espèces de mangroves recensées dans le monde se retrouvent en Outre-mer avec une répartition inégale : en effet seul 5 espèces sont présentes en Guyane, alors que la Nouvelle Calédonie en compte 26.
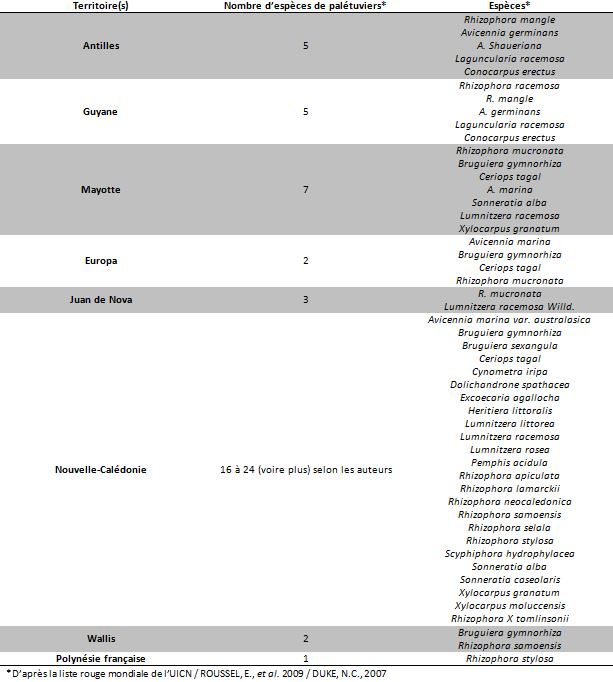
Les mangroves françaises représentent environ 0,6 % des surfaces de mangrove dans le monde (WCMC 2005). Sur le bassin Atlantique on les retrouve sur tous les territoires français de la zone intertropicale : à Saint Barthélémy, à Saint Martin, en Guadeloupe, en Martinique et en Guyane. Dans la zone Indopacifique on retrouve ces écosystèmes à Mayotte, aux îles Eparses, en Nouvelle Calédonie, à Wallis et Futuna et plus anecdotiquement en Polynésie où la présence de palétuviers est attestée sur l’île de Moorea et certains territoires des Îles sous le Vent. Il s’agit de l’espèce Rhizophora stylosa introduite à partir de 1933 à des fins ostréicoles (ROUSSEL et al. 2009). La surface des mangroves d’outre-mer a été estimée à 91 055 ha en 2016, répartie en majorité en Guyane et en Nouvelle Calédonie.
Forêt marécageuse
D’un point de vue géographique, la forêt marécageuse est la formation forestière qui fait suite à la mangrove dans les secteurs inondables (IMBERT et al. 1988). À la différence de cette dernière, la forêt marécageuse évolue dans des eaux peu ou pas salées. Elle se localise dans les fonds de vallée ou les plaines inondables littorales, toujours dans des sols hydromorphes. Les cortèges floristiques associés sont beaucoup plus diversifiés qu’en mangrove et diffèrent selon l’aire géographique.
Aux Antilles, la strate arborescente est dominée par une seule espèce, Pterocarpus officinalis, dont l’appellation vernaculaire diffère selon les localités (« Sangdragon » ou « Mangle médaille » en Guadeloupe, « Sangrillo » au Costa Rica, «Moutouchi-marécage » en Guyane…). La physionomie du fruit (en forme de médaille) ou la couleur rougeâtre de sa sève lui ont valu ces noms.


L’espèce est beaucoup plus rare en Martinique qu’en Guadeloupe. En Guyane, Pterocarpus officinalis côtoie d’autres espèces dominantes d’arbres de la forêt marécageuse : Euterpe oleracea, Virola surinamensis, Symphonia globulifera, etc. (CADAMURO, 1999). On appelle pinotière les forêts marécageuses guyanaises dominée par l’espèce Euterpe oleracea.

En Nouvelle-Calédonie, dans les dépressions marécageuses périodiquement inondées qui parsèment les plaines littorale, existe une formation spéciale : la forêt marécageuse à niaoulis. Il s’agit d’une forêt dense et fermée dont la strate arborée est représentée par une seule espèce : le niaouli (Melaleuca quinquenervia) qui, les pieds dans l’eau, trouve dans ce milieu son optimum écologique. Les sols sont hydromorphes, avec un horizon de gley en profondeur, et submergés la plus grande partie de l’année (Atlas de la Nouvelle-Calédonie – planche 15).


Marais saumâtres, vasières
Les marais littoraux, ou vasières, sont des écosystèmes caractérisés par la présence d’herbacées et d’un sol hydromorphe vaseux. Ils sont également sous l’influence de bon nombre de processus côtiers : marnage déterminant les paramètres de submersion, salinité de l’eau, mouvements sédimentaires… Quand elle est colonisée par la forêt, la vasière devient une mangrove.
Aux Antilles, les marais saumâtres sont le plus souvent situés entre la mangrove et la forêt marécageuse (IMBERT et al. 1988). La recolonisation de ces étendues par les espèces forestières des environs est également rendue difficile par la densité du couvert herbacé qui gêne la pénétration de leurs semences, celles-ci étant le plus souvent disséminées par flottaison. La lisière entre ces marais et la mangrove est souvent ouverte, comportant une végétation clairsemée. De tels endroits sont particulièrement propices pour les oiseaux d’eau et certains oiseaux marins (hérons, limicoles, poules d’eau…) qui y trouvent une nourriture abondante (IMBERT et al. 1988).
Marais intérieurs et savanes inondées de Guyane
Les marais intérieurs se développent sur des sols non salés et gorgés d’eau (IMBERT et al. 1988). Les cortèges faunistiques et floristiques associés diffèrent selon la localisation géographique des territoires d’Outre-mer. Ainsi, en Guadeloupe, la flore est caractérisée par la fougère Thelypteris interrupta, la graminée Echinochloa pyramidalis qui peut atteindre deux mètres de haut et former par endroits de véritables sols flottants qui ondulent sous les pas, l’ « Herbe couteau » Rhynchospora corymbosa qui rend le couvert végétal difficilement pénétrable et l’espèce ligneuse Chrysobalanus icaco. Ces espèces constituent alors des groupements purs ou mélangés que l’on rencontre principalement aux abords de la forêt marécageuse (IMBERT et al. 1988).
En Guyane, le terme de savane renvoie à un type de milieu humide ouvert et inondé, sorte de continuum écologique avec le marais. L’on distingue alors la savane sèche, la savane herbeuse inondable (inondée en saison des pluies et couverte de graminées, de cypéracées et d’arbustes tels que le poirier savane et l’oreille d’âne), la savane arbustive ou arborée composée d’herbes hautes et d’arbrisseaux comme les pruniers icaques et le poirier savane et enfin la savane inondée. Celle-ci, inondée toute l’année, porte également le nom local de « pripris », plus proche de l’écosystème de marais que celui de savane. Il s’agit de grandes étendues sur lesquelles poussent des herbes hautes, des palmiers bâche ainsi que des fougères associées au moucoumoucou (Montrichardia arborescens), une plante aquatique commune à la tige charnue et épineuse (CNES).


Mares
Le Pôle-relais mares, zones humides intérieures et vallées alluviales définit les mares comme « une étendue d’eau à renouvellement généralement limité, de taille variable et de 5000 m² au maximum. Sa faible profondeur qui peut atteindre environ deux mètres, permet à toutes les couches d’eau d’être sous l’action du rayonnement solaire, ainsi qu’aux plantes de s’enraciner sur tout le fond. De formation naturelle ou anthropique, elle se trouve dans des dépressions imperméables, en contextes rural, périurbain voire urbain. Alimentée par les eaux pluviales et parfois phréatiques, elle peut être associée à un système de fossés qui y pénètrent et en ressortent ; elle exerce alors un rôle tampon au ruissellement. Elle peut être sensible aux variations météorologiques et climatiques, et ainsi être temporaire. La mare constitue un écosystème au fonctionnement complexe, ouvert sur les écosystèmes voisins, qui présente à la fois une forte variabilité biologique et hydrologique interannuelle. Elle possède un fort potentiel biologique et une forte productivité potentielle » (http://www.pole-zhi.org/mare).


En Guadeloupe, le territoire de la Grande Terre est ponctué par de nombreuses mares. Mésestimées et menacées, nombre d’entre elles ont disparues dans les dernières décennies, victimes principalement des remblais pour l’aménagement du territoire. A l’échelle de l’Outre-mer tropical français, c’est l’une des principales menaces qui pèsent sur ces écosystèmes fragiles. Aux Abymes, le territoire des Grands Fonds comprend un réseau d’une soixantaine de mares, dernièrement recensées dans le cadre de l’élaboration du Plan local d’urbanisme.
Lacs et étangs
Selon le Pôle-relais mares, zones humides intérieures et vallées alluviales, un lac se "caractérise par une grande profondeur et une superficie importante permettant une stratification thermique de l’eau. Les bordures peuvent donc être colonisées par la végétation créant une zone humide, par opposition à la majorité de la surface appelée zone d’eau profonde. Les lacs peuvent être d’origine naturelle (verrous glaciaires, essentiellement en montagne) ou bien d’origine artificielle : retenues, pour stocker de l’eau à des fins de consommation, d’électricité, de régulation des crues. Les retenues sont sujettes à des variations importantes de niveaux d’eau (marnage) peu favorables à la végétation aquatique et amphibie. En revanche, la mise à découvert de vasières en fin d’été attire les oiseaux sur leur voie migratoire. La préservation et le développement durable de ces zones humides relèvent d’enjeux comparables à ceux des zones d’étangs et marais mais le plus souvent, à une plus grande échelle avec les pollutions des eaux liées à l’urbanisation et l’industrialisation, à l’agriculture intensive, à la fréquentation humaine des berges. Ainsi que la dégradation et/ou disparition des zones humides de proximité sous les mêmes pressions."
Un étang se distingue du lac de par sa profondeur et sa superficie de moindre importances.


Différentes conditions amènent à la création de plans d’eau d’altitude d’origine volcanique. Ceux-ci peuvent occuper l’ancienne caldeira d’un volcan, il s’agit alors de lacs ou d’étangs de cratères. On peut notamment évoquer le lac Dziani Dzaha à Mayotte, ou encore le Grand-Étang de La Réunion, unique étang d’altitude d’origine volcanique de l’île. Ce sont des écosystèmes particuliers, dont les processus chimiques et biologiques peuvent différer de ceux que l’on retrouve dans les lacs et étangs situées en zone de plaine. Une récente étude scientifique du lac Dziani Dzaha le présente comme un écosystème unique où les cyanobactéries conditionnent le cycle du carbone et la biomasse phytoplanctonique dans un milieu ressemblant davantage à celui de l’eau de mer. Le territoire de Wallis-et-Futuna compte jusqu’à 7 lacs de cratères (lacs Lalolalo, Lanu, Lanutavake, Kikila, Lanutuli, Alofivai, Lanumaha).
Tourbières
Les tourbières sont des écosystèmes caractérisés par la présence d’un sol hydromorphe, empêchant la pénétration de l’oxygène : c’est un milieu anaérobique qui limite fortement la présence des microorganismes responsables de la décomposition de la matière organique. Celle-ci n’est donc que partiellement recyclée et finit par former un dépôt noirâtre que l’on nomme « la tourbe ».
Dans l’outre-mer tropical, on retrouve notamment ces types d’écosystème en Guyane (marais de Kaw, Mana) et à la Réunion (tourbière du plateau de Thym, zones humides du Piton dans l’Bout, Piton de la Fournaise…).
Récifs coralliens
Les récifs coralliens et les fonds marins inférieurs ou égaux à 6 m de profondeur sont inclus dans la définition Ramsar des zones humides. Les récifs coralliens résultent de la présence d’êtres vivants, ce qui constitue leur originalité. Les coraux sont des animaux qui vivent en symbiose avec une algue (apport d’oxygène via la photosynthèse) qui constitue leur squelette calcifié et rigide. Les madréporaires, aux polypiers constitués de carbonate de calcium (96 à 99,70 %), représentent l’armature des récifs.
On distingue plusieurs types de récifs coralliens : les récifs frangeant, qui bordent étroitement la côte comme aux Antilles, les récifs barrières en bordure extérieure d’un lagon (cas de la Nouvelle Calédonie, de la Réunion), les bancs récifaux construit au large sur des hauts fonds (Grande barrière d’Australie fixée sur le rebord d’une plateforme continentale) et les atolls, véritables couronnes de calcaire récifal entourant des lagons sans îles rocheuses centrales mais parfois parsemés d’îlots coralliens (PASKOFF, 2004).
L’écologie des récifs coralliens est en interrelation avec d’autres types d’écosystèmes, terrestres et marins. On parle alors d’écosystèmes associés : tout cet ensemble doit être appréhendé à travers un continuum écologique terre-mer. Par exemple, les mangroves participent à piéger les sédiments issus de l’érosion limitant ainsi la turbidité de l’eau. Une turbidité trop élevée peut altérer le processus de photosynthèse des algues symbiotiques et donc nuire à la bonne santé des récifs. Les herbiers de phanérogames font également partis des écosystèmes associés dans la mesure où ils favorisent également la retenue de sédiments et l’oxygénation de l’eau.
Prairies humides
Les prairies humides se développent sur les terrasses alluviales humides, à proximité de cours d’eau lents, ou à l’occasion de replats détrempés parfois parcourus par des ruisseaux.
Elles sont constituées de formations végétales denses, de hauteur moyenne à assez haute qui peuvent atteindre 1,50 m. Cette végétation est dominée par des graminées sociales ou des dicotylédones coloniales (Pôle-relais zones humides intérieures). En Guadeloupe, les prairies humides d’arrière mangrove se développent sur des sols argileux très compacts et salés : la diversité des espèces floristiques y est alors limitée. Il s’agit essentiellement de l’Amaranthe bord-de-mer (Philoxerus vermicularis) et de la graminée Sporobolus virginicus. Dans les secteurs peu ou non salés, les prairies présentent des cortèges floristiques plus riches avec notamment des Cypéracées (genres Cyperus, Eleocharis et Fimbristylis), des Graminées et des Papilionacées. Trois espèces de limicoles migrateurs dépendent directement de ces étendues herbacées marécageuses : le pluvier doré (Pluvialis dominica), la Poule vergenne (Bartramia longicauda) et la Bécassine (Capella delicata) (IMBERT et al. 1988).
Lagune
On parle de lagune pour désigner un plan d’eau littoral plus ou moins saumâtre séparé de la mer par un cordon sableux. Les échanges avec le milieu marin sont plus ou moins affirmés selon la configuration de la lagune, plus ou moins fermée. Lorsqu’un récif corallien sépare une étendue d’eau vis-à-vis de la mer, on parle alors de lagon.
Plaine alluviale
Une plaine alluviale se caractérise de par sa topographie plane ou quasi-plane. Située en zone de thalweig, elle est constituée d’alluvions soit les produits de l’érosion et des divers matériaux charriés par les cours d’eau. La plaine alluviale correspond à un secteur inondée lors des crues ou de la saison humide.
L’une des entités caractéristiques de plaine alluviale dans l’outre-mer tropical français est celle de la plaine alluviale de Kaw, en Guyane.
Sources
CADAMURO, L. 1999. Structure et dynamique des écosystèmes inondables (forêt marécageuse, mangrove) du bassin Sinnamary (Guyane française). Thèse : Écologie tropicale. Université Paul-Sabatier – Toulouse III. 255 p.
DUKE, N.C. 2007. A brief account of the mangroves of New-Caledonia - Interim rapport. University of Queensland, QLD Australia. 4 p.
IMBERT, D. et al. 1988. Les milieux humides du littoral guadeloupéen. Office national des forêts. 61 p.
PASKOFF, R. 2004. Côte en danger. L’harmattan. Paris. 247 p.
ROUSSEL, E. et al. 2009. Les mangroves de l’outre-mer français ; Écosystèmes associés aux récifs coralliens. Documentation Ifrecor. 145 p.
VIRLY, S. 2008. Atlas des mangroves de Nouvelle-Calédonie. ZoNéCo. 208 p.